По прошествии 31 000 поколений в одной из 12 популяций (ее условное обозначение – Ara-3) произошло нечто странное. Ее численность теперь продолжала расти и после того, как вся глюкоза в колбе была съедена. К концу суточного цикла численность популяции Ara-3 оказывалась намного выше, чем у остальных популяций. Очевидно, микробы научились использовать в пищу какое-то вещество, входящее в состав среды, но несъедобное для обычных кишечных палочек.
Вскоре Ленски и его коллеги выяснили, что бактерии из популяции Ara-3 приобрели способность питаться цитратом (лимонной кислотой). Цитрат добавляется в среду в качестве вспомогательного вещества (хелатирующего агента). Нормальные бактерии E. coli способны усваивать цитрат только в анаэробных условиях. Неспособность питаться цитратом в присутствии кислорода – один из определительных признаков данного вида бактерий. Таким образом, в популяции Ara-3 появилось настоящее «эволюционное новшество» – новый полезный признак (авторы обозначили его Cit+), изменивший взаимоотношения организма со средой и позволивший бактериям-мутантам увеличить свою численность и стать доминирующими в популяции. Любопытно, что бактерии Cit+ так и не вытеснили до конца бактерий Cit–, которые питаются только глюкозой. Возможно, их длительное мирное сосуществование объясняется разделением ниш, но это пока лишь гипотеза.
Дизайн долгосрочного эксперимента позволяет проследить формирование эволюционного новшества во всех генетических подробностях. Именно для решения подобных задач эксперимент и был поставлен. Часть представителей каждой популяции периодически замораживают. Это нисколько не вредит здоровью микробов. В любой момент можно их оттаять и использовать для повторных опытов. Это, в частности, позволяет оценить вероятность того или иного эволюционного события: насколько оно было случайным или закономерным.
Авторы отсеквенировали геномы 29 бактерий из популяции Ara-3, замороженных в разное время. Это позволило построить эволюционное дерево, показанное на рис. 1. Оказалось, что популяция оставалась генетически разнообразной на протяжении почти всей своей истории. В результате мутаций и отбора из одного предкового клона получилось несколько генетически различающихся линий (клад), но они не спешили вытеснять друг друга. Скорее всего, это значит, что большинство генетических различий между кладами были нейтральными, то есть не влияли на приспособленность (а в некоторых случаях, как уже говорилось, могло идти взаимное приспособление и раздел ниш).
Исследователи выявили конкретное генетическое изменение, которое дало бактериям возможность питаться цитратом. Микробы Cit– превратились в Cit+ благодаря дупликации (удвоению) фрагмента хромосомы, содержащего ген citT (рис. 2). Этот ген кодирует белок, транспортирующий цитрат из внешней среды в цитоплазму бактериальной клетки. В норме у E. coli ген citT активен только в анаэробных условиях. Однако в результате дупликации одна из двух копий citT попала под управление промотора другого, соседнего гена (rnk). При помощи генно-инженерных экспериментов авторы показали, что промотор гена rnk обеспечивает работу контролируемого им гена в аэробных условиях.
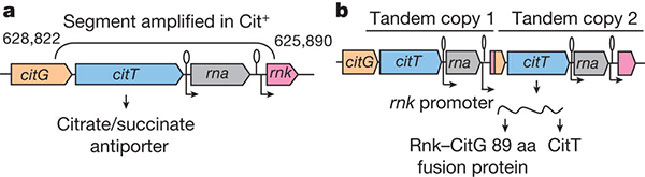
Рис. 2. Тандемная дупликация, которая привела к появлению способности питаться цитратом у бактерий из популяции Ara-3. а — участок хромосомы предкового штамма, b — тот же участок после дупликации. Стрелками и петельками обозначены промоторы генов rna и rnk, обеспечивающие работу этих генов в аэробных условиях. В результате дупликации ген citT, кодирующий транспортер цитрата, оказался под управлением промотора гена rnk и стал экспрессироваться в присутствии кислорода. Рисунок из обсуждаемой статьи в Nature
Разобравшись с природой ключевой мутации, авторы задались следующим вопросом: было ли появление фенотипа Cit+ в кладе 3 популяции Ara-3 полностью случайным? Могло ли это событие с тем же успехом произойти в другой кладе или в другой популяции, или оно было подготовлено предшествующей эволюцией именно этой клады? В поисках ответа авторы провели повторные эволюционные эксперименты с размороженными представителями клад 1, 2 и 3, а также с исходным (предковым) штаммом.
Выяснилось, что у предкового штамма вероятность появления фенотипа Cit+ пренебрежимо мала. Представители клады 1 выработали этот фенотип в двух повторных экспериментах из 55. Микробы из клады 2 научились питаться цитратом в двух случаях из 97. Наконец, бактерии из клады 3 приобрели эту способность в восьми случаях из 37 повторных опытов.
Приобретение фенотипа Cit+ во всех случаях было связано с тем, что ген citT начинал экспрессироваться в аэробных условиях. Но достигался этот результат разными путями. В части повторных опытов, как и в основном долгосрочном эксперименте, произошли тандемные дупликации, благодаря которым citT оказался под управлением аэробного промотора. Но границы дуплицированных участков в разных случаях были разными, а промотор, взявший на себя управление геном citT, не всегда был промотором гена rnk: в одном случае это был промотор гена rna. В нескольких повторных опытах тот же эффект был достигнут без дупликации – благодаря встраиванию мобильного генетического элемента IS3 в кодирующую часть гена citG, расположенного по соседству с citT (на рисунке слева). В этом мобильном элементе как раз имеется подходящий промотор.
Повторные эксперименты показали, что вероятность приобретения полезного признака Cit+ постепенно росла в ряду «предковый штамм – клады 1 и 2 – клада 3». Стало быть, появление данного признака именно в кладе 3 не было простой случайностью. Микробы были заранее подготовлены к этому – «потенцированы». Иными словами, у них в ходе предшествующей эволюции зафиксировались какие-то мутации, которые сами по себе не давали возможности питаться цитратом, но повысили вероятность развития этой способности в будущем. Разумеется, «потенцирующие» мутации зафиксировались не потому, что микробы планируют свою эволюцию наперед. Эти мутации либо были нейтральными и зафиксировались случайно, либо были полезны для чего-то другого. Подготовка к приобретению способности питаться цитратом была побочным эффектом, которого естественный отбор не мог предусмотреть.